1Department of Gastrointestinal Surgery, Xiang’an Hospital of Xiamen University, School of Medicine, Xiamen University, Xiamen, Fujian 361000 China
2Department of Gastroenterology, Xiang’an Hospital of Xiamen University, School of Medicine, Xiamen University, Xiamen, Fujian 361000 China
3Department of Gastrointestinal Surgery, The First Affiliated Hospital of Sun Yat-sen University, Sun Yat-sen University, Guangzhou, Guangdong 510080 China
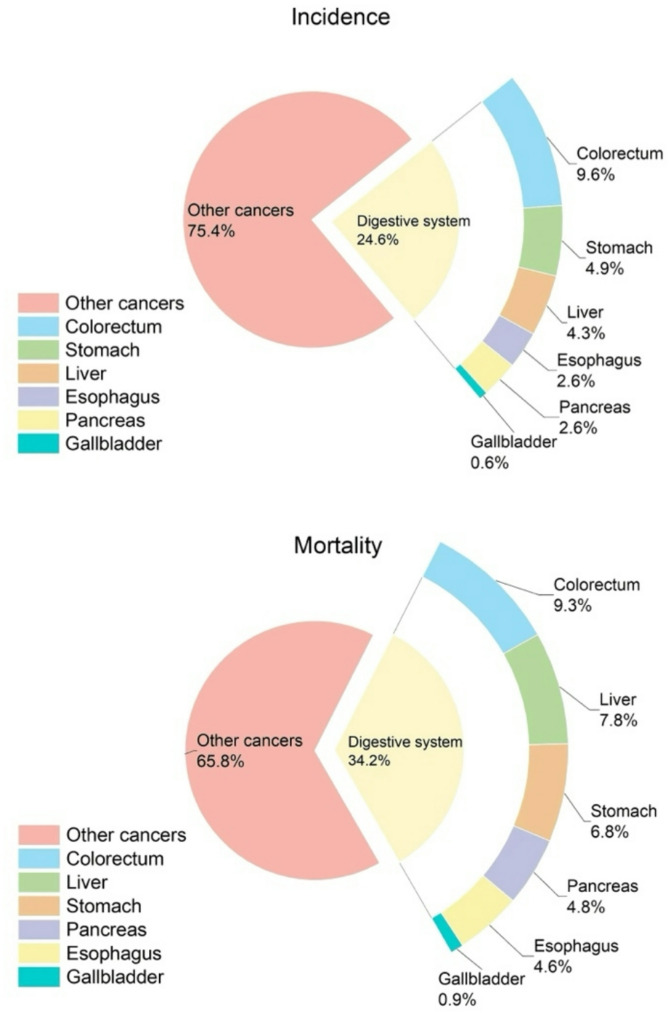
1]and the trend of incidence and mortality has been increasing year by year, which is a serious risk to people’s health (Fig. 1).
Incidence and mortality of colorectal cancer and other digestive system tumors. Colorectal cancer is the third most common cancer in the world and the second most common cause of cancer-related deaths
Although innovations in molecular biology techniques and tumor treatment strategies have significantly advanced the field of cancer research, tumor metastasis remains a central causative factor in the deaths of cancer patients worldwide [2]leading to approximately 90% of cancer-related deaths. Tumor metastasis also dominates the high mortality rate of cancers of the digestive system such as colorectal cancer. Once a primary tumor, which could have been intervened with a variety of methods such as surgery, chemotherapy, radiotherapy, targeted therapy and immunotherapy, has progressed to the metastatic stage, the options for clinical intervention options are significantly limited.
In this context, the study of circulating tumor cells (CTCs), as a key mediator of tumor metastasis, has become an important breakthrough in revealing metastatic mechanisms and developing intervention strategies. Circulating tumor cells are tumor cells that enter the blood circulation after detachment from primary and/or metastatic tumors, and they carry key tumor biological information through haematogenous dissemination and eventually colonize the distal organs to form metastatic foci. Since the first discovery of CTCs in the blood of a patient with metastatic cancer by Thomas Ashworth in 1869 [3], CTCs have gradually gained prominence in the fields of tumor biology, molecular analysis and precision medicine.
The metastatic pathways of tumors are extremely complex. A full understanding of the mechanism of CTC in metastasis of digestive system tumors such as colorectal cancer and the interactions in the infiltration-metastasis cascade response is crucial in multiple dimensions such as prevention and early diagnosis of tumor metastasis, and identification of new potential therapeutic targets.
In this review, the biological properties of CTC and its relevant mechanisms for inducing metastasis in digestive system tumors are reviewed. In addition, CTC-related preclinical models and numerous CTC enrichment and isolation techniques are briefly summarised. Finally, we also retrospectively analyse the diagnosis and treatment of CTC in tumors such as CRC and discuss the great potential of CTC in clinical applications.
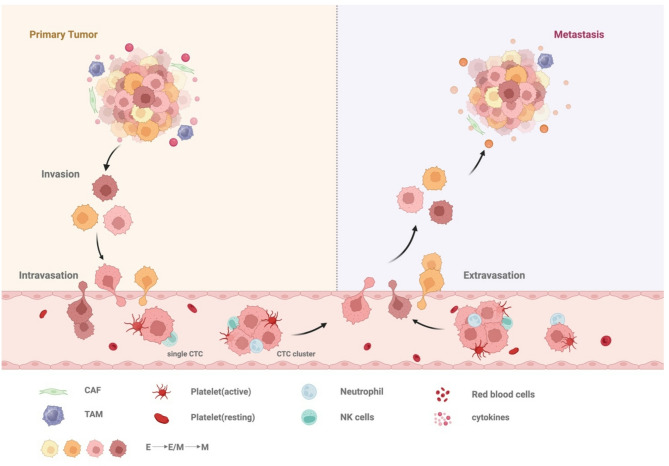
2). In colorectal cancer liver metastasis (CRLM), for example, CRC cells need to change the tumor microenvironment (TME) first, and in the initiation phase of the invasion-metastasis cascade reaction, colorectal cancer (CRC) cells inhibit the expression of E-cadherin and disrupt the junctional structure between epithelial cells to detach from the primary site [4]. Subsequently, secretion of matrix metalloproteinases (MMPs) and activation of urokinase-type plasminogen activator (u-PA) lead to degradation of the basement membrane. At the same time, pro-angiogenic factors (e.g., VEGF) released by ECM degradation can induce neovascularisation and lymphangiogenesis, providing a gateway for CTCs to enter the circulatory system [5].
CTC-mediated tumor metastatic process. The process of tumor metastasis is very complex, and can be roughly divided into tumor cells from the primary tumor site intravasation into the blood circulation to become CTCs, CTCs interacting with blood cells and so on in the circulation, and some of the surviving CTCs undergoing extravasation and ultimately achieving colonization in distant locations
Upon entering the circulation, CTCs enhance their own resistance to blood flow shear forces by forming adhesion complexes with other blood cells and inhibit loss-of-nest apoptosis. Notably, only a small number of circulating tumor cells are able to adapt to the microenvironmental pressure of the target organ and evade immune clearance, seeking out the hostile environment of the circulation and eventually extravasating to the liver, where they successfully colonise pre-metastatic ecological niches that have been reprogrammed in advance [6]. Moreover, CTCs arriving at the target organ may enter a dormant state with cell cycle arrest and activation of pro-survival signalling pathways.
How CTCs originated
Cellular infiltration of tumor cells and intravasation into adjacent blood vessels is the first step in the development of metastasis [7] (Table 1).
Table 1.
Key mechanisms and pathways of CTC-mediated tumor metastasis
Invasion-metastasis cascade
Mechanisms
Key molecules/pathways
References
How CTCs originated
EMT
- EMT enhances invasiveness and metastatic potential;
Epithelial-mesenchymal transition (EMT) is the initiating step of metastasis [8–10] and refers to the reversible process of transient transdifferentiation of epithelial cells into motile mesenchymal cells [11] (Fig. 3).Tumor cells are endowed with enhanced invasiveness and metastatic potential after the EMT process, which in turn induces tumor cell intravasation and detachment [7, 12]. This process occurs by the orchestration of EMT-inducible transcription factors (EMT-TFs). core EMT-TFs such as SNAIL family members (Snail, Slug), TWIST family (TWIST1 and TWIST2), and E-box-binding (ZEB) transcription factors [13, 14]in a variety of combinations, lead to epithelial phenotypic repression and mesenchymal phenotypic activation of gene expression changes [15–18]. Among them, matrix metalloproteinases (MMP) of the zinc-dependent endopeptidase family can be activated by Snail and Zeb2 to degrade ECM components [11, 19] for tumor cell invasion. In addition to this, it has been shown that ZEB1 is positively correlated with cancer invasiveness, that mutations in the Transcriptional repressor GATA binding 1 (TRPS1) can directly bind to the ZEB1 promoter to activate the onset of transcription, and that the TRPS1 R544Q/ZEB1 axis is closely associated with CRC metastasis, which ultimately leads to a poor prognosis for CRC patients [20]. However, another study suggests that ZEB1 can paradoxically inhibit the EMT process in BRAF-mutated CRC, presenting a role as a tumor suppressor [21].
EMT and MET. Epithelial-mesenchymal transition is the initiating step of metastasis. Epithelial-mesenchymal transition (EMT) drives tumor cells to leave their primary foci and enter the circulatory system by conferring them with the ability to migrate and invade, while mesenchymal-epithelial transition (MET) encourages disseminated tumor cells to regain epithelial properties and colonize the distal organs to form metastatic foci. This dynamic and reversible EMT-MET transition synergistically regulates the whole process of tumor metastasis, including invasion-dissemination-colonization
Tumor-associated macrophages (TAMs) also play a facilitating role in EMT formation. Both HCC cells and pancreatic cancer cells stimulate TAMs to undergo M2 polarization, which promotes the progression of EMT [22, 23]. In the process of colorectal cancer liver metastasis, tumor-associated macrophages (TAMs) and mesenchymal phenotypic tumor cells form a vicious circle through the bidirectional regulation mechanism of IL-6/STAT3-CCL2 axis, which ultimately promotes the formation and progression of liver metastases [24]. Besides, TAMs are involved in almost the whole process of tumor metastasis. Recent studies have shown that through comparative analysis of primary foci and paired liver metastatic tissues from patients with primary colorectal cancer, researchers identified a Subpopulation of TAMs characterized by high expression of secreted phosphoprotein 1 (SPP1), which was significantly correlated with enhanced tumor aggressiveness and reduced patient survival [25] (Table 2).
Table 2.
Roles of immune cells in regulating CTCs across the metastatic cascade
Cell type
Interactions
Key molecules/pathways
References
Neutrophils/NETs
- Induces angiogenesis and intravasation
- Promotes extravasation and metastasis formation
- Enhances endothelial adhesion
- Inhibits immune cell attacks on tumor cells
MMP9, ICAM-1/Mac-1, β1-integrin, IL-17, IL-1β, matrix metalloproteinases, etc.
Similarly, exosomes are also an important part. EVs can enhance the shedding ability of CTCs by carrying EMT-promoting factors (e.g., TGF-β) and regulating key genes in the EMT pathway [26]. It has been suggested that exosomal long chain non-coding RNA (lncRNA) PCAT1 can induce EMT, which in turn promotes CTC migration leading to colorectal cancer liver metastasis [27].
In addition, research results indicate that gut microbiota can also influence the EMT process. Research findings indicate that F. nucleatum reduces E-cadherin expression in colorectal cancer cells and increases vimentin expression, suggesting that F. nucleatum may promote colorectal cancer metastasis by influencing the epithelial-mesenchymal transition (EMT) process [96].
Interestingly, EMT does not result in a single mesenchymal state; cells are usually in “epithelial-mesenchymal plasticity” (EMP). In this case, the cells are in various intermediate states with varying degrees of epithelial and mesenchymal characteristics, rather than a simple binary switching pattern. The epithelial state is retained both because the high proliferation rate associated with the epithelial phenotype is essential for colonization of distant organs and because a complete transition to a mesenchymal phenotype would limit essential protein interactions [97]. Among them, E- and E/m-type cells have enhanced adhesion and extravasation capacity, which facilitates proliferation and metastasis, while M/e and M-type cells are associated with tumor recurrence [98]. One study showed that the ratio of m-CTCs/e-CTCs was significantly higher in patients with postoperative recurrence of pancreatic cancer than in patients without recurrence (0.23:0.1) [99]. In addition, the m-CTCs to ratio was a negative factor for postoperative HCC recurrence [100].
Vascular damage
During colorectal cancer metastasis, tumor cells undergoing EMT can target and inhibit the expression of VE-calmodulin (VE-Cad) and p120-catenin in vascular endothelial cells through the release of exosomes rich in miR-27b-3p, disrupting endothelial cell tight junctions and leading to the loss of vascular barrier integrity [28]. This abnormal elevation of vascular permeability aims to significantly increase the generation and dissemination of circulating tumor cells by remodeling the microenvironment at the tumor-vessel interface, forming a cascade of “EMT-exosomes-vascular leakage-CTC release” to promote metastasis.
Data from another study showed a significant positive correlation between the level of ADAM17-positive exosomes and the number of CTCs in the peripheral blood of colorectal cancer patients. Explained at the mechanistic level, ADAM17 specifically shears VE-calmodulin in vascular endothelial cells, leading to disruption of endothelial barrier integrity and increased vascular permeability [29]. This alteration provides the microenvironmental basis for the formation of pre-metastatic niche and ultimately leads to distal organ colonization of colorectal cancer via the blood pathway.
How CTCs survive in the bloodstream
In the circulation, CTCs will face many challenges such as shear stress, immune system attack, and so on, but the formation of CTC clusters and hypoxic microenvironment can protect some CTCs from surviving in the circulation for extravasation and eventual metastasis [101] (Table 1; Fig. 4).
CTC in the blood microenvironment. CTC interact with neutrophils, platelets, CAFs, and TAMs, among others, in the circulation to resist many challenges including shear stress, immune system attack, and loss-of-nest apoptosis
Surviving shear stress
Some of the CTCs entering the arteries may fragment and die due to high shear stress [102]thus regions of low shear stress are more susceptible to attack by tumor cells and thus intravasation. During circulation, integrins contribute to CTC-dependent adhesion to low shear stress regions of the vessel wall. It has been shown that β1 integrin and CD44 can help CTC resist shear forces and save them from detachment from the endothelial cell layer [30–32]. It has been shown that Talin-1, an adhesion plaque protein present in integrin-associated complexes, helps to activate integrin β1, which promotes transendothelial migration and subsequent formation of liver metastases in colon cancer cells [33] (Table 2).
Ion mobility is also thought to be associated with the resistance of CTC to shear stress. CTC protects the plasma membrane from damage in the face of shear stress by increasing cellular stiffness, which is associated with Ca²⁺ and Na⁺/H⁺ exchange [34].
The unique structural characteristics of CTC clusters result in a much lower hydrodynamic resistance to their movement in the bloodstream. It has been shown that although CTC clusters are less numerous than individual CTCs in the circulation, their metastatic potential is 23–50 times higher than the latter [98]which may be related to the fact that CTC clusters up-regulate cellular connectivity components, such as spotted bead proteins, and the associated blood cell crosstalk [35]. It has been shown that tumor cells undergo membrane damage due to shear stress in the absence of physical protection from platelets [36]. When CTCs express tissue factor on their surface, platelets are activated and recruited to form a “platelet microthrombus” around CTCs, which physically protects CTCs under the threat of shear stress. In addition, this protective clot also attracts monocytes/macrophages to recruit to the CTC [37, 38]helping the CTC to adhere to the endothelium and undergo further extravasation (Table 2). Similarly, CAF is resistant to high shear. In addition, it has been shown that intra-CTC bacteria can regulate the cytoskeleton to undergo rearrangement and enhance the viability of CTCs under mechanical stress in order to improve the survival of CTCs in circulation, correlating with the mechanism by which cytoplasmic bacteria inhibit RhoA and ROCK activation [39].
Resisting attacks from immune system
The blood environment is not friendly to CTC [103]but at the same time a number of components are used by CTC for their metastatic purposes (Table 2).
Platelets are involved in almost the entire process of tumor metastasis as “accomplices” [104]. TGF-β secreted by platelets can activate Smad and NF-κB signaling pathways in tumor cells [40] in order to induce the EMT state of CTC [41] and enhance the invasive ability of CTC. In addition, platelets can activate the FAK/JNK/c-Jun signaling axis in CTC through physical contact and transcriptionally up-regulate the immunosuppressive checkpoint molecule CD155, which confers the ability of CTC to escape NK cell killing and promotes distant metastasis of hepatocellular carcinoma [42]. Platelets may also exhibit the ability to promote CTC extravasation at later stages of tumor cell metastasis. The resistance of platelets to immune surveillance might serve as a future chemotherapeutic target for tumor metastasis [105]. Meanwhile, with the breakthrough of nanomedicine, the development prospect of platelet membrane for drug delivery in cancer therapy is also very promising [106].
NK cells could have mediated lysis and thus recognition and elimination of CTCs to prevent further progression of tumor metastasis, but the interaction of NK cell receptor D with the MICA/MICB of CTCs prevents this process from occurring, thus giving CTCs an opportunity to take advantage of it [105].
For assisting CTC metastasis, the credit of neutrophils should not be underestimated as well. It has been shown that neutrophils can account for about 90% of the total WBCs that escort CTCs to metastasis in mouse models and up to 75% in patients [43]. Neutrophils can assist CTC in immune escape and significantly enhance their metastasis-related biological properties through two modes of interactions: (i) CTCs directly adhere to the top of neutrophils and hitchhike, and (ii) they indirectly bind to neutrophils through platelet-constructed molecular bridges. This dynamic interaction network not only stabilizes the mechanical integrity of CTC, but also enhances its vascular wall anchoring and transendothelial migration by activating adhesion signaling pathways. Notably, CTC-neutrophil clusters are significantly superior to isolated CTCs in terms of metastatic potential, with a significant positive correlation between their blood detection rate and the invasive progression of malignant tumors such as breast, gastric, and hepatocellular cancers, and the presence of CTC-neutrophil clusters is associated with a worse prognosis for patients [44]. Therefore CTC-neutrophil cluster has the potential to be an important biomarker for assessing the risk of tumor metastasis. Neutrophils form adhesive meshwork structures called neutrophil extracellular traps (NETs), which can protect CTCs from NK cell-mediated cytotoxicity by forming physical barriers and secreting immunosuppressive cytokines.
Preventing anoikis
Loss-of-nest apoptosis is a protective barrier against metastatic cell dissemination. After detaching from extracellular matrix (ECM) anchorage, circulating tumor cells need to overcome loss-of-nest apoptosis (anoikis) - a specific process of programmed death directly induced by the disruption of intercellular adhesion or extracellular matrix attachment [107]. Normal epithelial cells activate adhesion spot kinase (FAK) via integrins and initiate the PI3K/AKT and RAS-RAF-MEK-ERK signaling pathways [108]. When anchorage deficiency occurs, metabolic disorders such as activation of the mitochondrial apoptotic pathway [109] or ATP depletion can trigger cell death [110]. However, tumor cells remodel the survival network through multidimensional mechanisms such as endogenous signaling reprogramming, microenvironmental adaptive remodeling, and metabolic stress compensation to compensate for integrin signaling deficits, inhibit mitochondrial translocation and phosphorylation of pro-apoptotic proteins, establish a pre-metastatic survival microenvironment, and scavenge reactive oxygen species to maintain energy homeostasis.
It has been shown that CPT1A-mediated activation of fatty acid oxidation (FAO) induces ROS scavenging and thus the ability of CRC cells to resist nest loss apoptosis [45]. In addition, TCF7L2 is highly expressed in GC as an important transcriptional regulator of urokinase-type plasminogen activator receptor (PLAUR) and promotes loss-of-nest apoptosis resistance and metastasis in GC cells [46]. It is noteworthy that viruses can also modulate anoikis resistance (AR) related pathways to maintain infected cell survival and promote metastasis [111]. For example, HBx proteins can drive hepatocellular carcinoma cells to bypass resistance to anoikis apoptosis [112] and promote hepatocellular carcinoma metastasis through multiple pathways synergistically (PI3K/AKT, EMT, PAK1, NF-κB and miRNA network).
Hypoxic microenvironment
A hypoxic environment can place tumors under a powerful selective pressure that not only drives malignant cell proliferation but also diminishes their susceptibility to apoptosis [47]. HIF signaling makes tumor cells more aggressive in a hypoxic environment [35]. It has been shown that overexpression of HIFs can promote tumor cells to undergo metastasis [48]and it has been suggested that it may be related to the ROS-resistant phenotype [35]. In addition to this, the aggregation of tumor cells can in turn induce HIF-1α-mediated mitochondrial autophagy and scavenge ROS [49]. Elevated levels of HIF-1α have been shown to correlate with poor prognosis in patients with gastrointestinal tumors such as colorectal cancer, gastric cancer, and hepatocellular carcinoma [50–52].
CTC clusters and microemboli
CTC clusters, although fewer in circulation than individual CTCs, have 23–50 times higher metastatic potential [53, 54] and better survival than individual CTCs [55, 56]. This is due to the own characteristics of CTC clusters. First, CTCs traveling in clusters seem to prefer to be released from hypoxic regions of the tumor [57]and their own cluster structure provides a hypoxic microenvironment [55]which facilitates their survival. Second, compared with individual CTCs, CTC clusters exhibit specific hypomethylation patterns associated with stem cell characteristics [58]such as OCT4, NANOG, SOX2, and SIN3A [59]which establishes CTC clusters to be more invasive and is closely associated with poorer prognosis. In addition, the residence time of CTC clusters in the circulation is much less than that of individual CTCs, which is conducive to the dissemination and colonization of tumor cells to distant organs, thanks to the characteristics of CTC clusters, whose own larger structure leads to slower mobility in the circulation and easier adhesion to the vessel wall [113].
In addition to homotypic CTC clusters formed between CTCs, CTCs can form heterotypic CTC clusters with other cell types such as neutrophils, myeloid-derived suppressor cells, platelets, and CAFs [114] or circulating tumor microemboli (CTM), which have the ability to resist shear stress, avoid anoikis [115] and immune escape [40]. It has been shown that patients with detectable CTC-neutrophil clusters have a poor prognosis [114]. Another study showed that MDSCs can help CTCs to be invisible in immune surveillance [116] and thus protected from the immune system (Table 2). In addition to this, tumor microemboli formed by aggregation of multiple cells are more likely to linger in the microvasculature of distal organs and form a pre-metastatic microenvironment by invading the vessel wall, as compared to single circulating tumor cells [117]. In addition to this, CD44-mediated signaling pathways are also responsible for the high efficiency of CTC cluster transfer [118].
Notably, medical manipulations such as puncture biopsy may also result in passive release of CTC from the primary tumor and portend poorer progression-free survival [119].
How CTCs form metastases
CTCs that have survived multiple challenges in the circulation next face extravasation and colonization. Even if they reach distant organs successfully, some CTCs may become dormant rather than forming metastases immediately (Table 1).
Extravasation
After overcoming multiple obstacles, some of the surviving colorectal cancer CTCs will be specifically captured into the hepatic microvascular system, and subsequently successfully infiltrate into the hepatic parenchymal microenvironment by interacting with endothelial cells and immune cells in the local capillaries, which ultimately mediate their transendothelial migration into the hepatic parenchyma (i.e., extravasation).
The main mechanisms of extravasation are paracellular migration (dominant) and transcellular migration (rare). It is noteworthy that host blood cells often play an “accomplice” role in the extravasation process. Platelets can enhance tumor cell invasiveness [40]and can also help CTCs to achieve extravasation in other direct or indirect ways, such as by inducing the opening of the endothelial barrier [60]. Neutrophils capture CTCs and enhance their endothelial adhesion by releasing NET [61]and can also promote CTC extravasation by releasing proteases or ROS to degrade ECM and increase vascular permeability. For colorectal cancer cells such as HT-29, CTC capture can be mediated by NET-associated CEACAM1 [62]. VEGF released by metastasis-associated macrophages (MAMs) can induce an increase in vascular permeability [63]which promotes CTCs extravasation (Table 2).
It has been shown that colorectal cancer cells can derive miR-25-3p after uptake by liver sinusoidal endothelial cells (LSEC), which leads to upregulation of VEGFR2 and enhanced vascular permeability [64]. Similar expression changes promote endothelial windowing, which further promotes extravasation.
Colonization
It is worth mentioning that CTC does not randomly colonize any organ and metastasize but shows organ tropism. When CTC leaves the primary tumor, its destination has already been predetermined and a supportive pre-metastatic niche (PMN) has already been formed.
Pre-metastatic niche formation
There is increasing evidence that primary tumors have released relevant factors including tumor-derived secretory factors (TDSF) and extracellular vesicles (EV) to the designated sites before CTCs reach the metastatic foci [65, 66]and these factors subsequently act on bone marrow-derived dendritic cells (BMDCs) or tissue-resident cells to reprogram the microenvironment of the distant target organ to make it suitable for CTC survival and colonization. Suitable “soil” prepared before the arrival of the “seed” seems to suggest that the treatment of tumor metastasis should focus on both the “seed” and the “soil” [67].
Tumor-derived secreted factors (TDSF): Previous studies have shown that TGF-β promotes colorectal cancer metastasis by recruiting tumor-associated macrophages (TAMs) and regulatory T cells (Tregs) and inducing the release of S100A8/S100A9 to drive epithelial-mesenchymal transition [68, 69]while IL-10 inhibits CD8 + T cells by up-regulating PD-L1 expression on monocytes infiltration and its mediated anti-tumor immune response, synergistically enhancing the process of colorectal cancer metastasis [70] (Table 2).
Extracellular vesicles (EV): EV can determine the site of CTC stagnation in the vascular system and can also assist in the development of extravasation [71]. Studies have shown that miR-934 and miR-203a-3p promote colorectal cancer metastasis by inducing M2 macrophage polarization to promote CXCL13 secretion and activation of the CXCL12/CXCR4/NF-κB pathway to drive the formation of pre-metastatic ecological niches, respectively [72, 73]; miR-181a-5p, on the other hand, activates a positive feedback loop through CCL20 secreted by hepatic stellate cells, thereby inducing tumor microenvironment (TME) reprogramming [74]; miR-135a-5p can initiate the LATS2-YAP-MMP7 signaling axis [75]; ADAM17 induces leaky angiogenesis by targeting vascular endothelial cells and accelerates the pre-metastatic ecological niche establishment [29]. Another study showed that CRC-derived extracellular vesicles can alter their gene expression through specific fusion with hepatic KCs, which in turn promotes pre-metastatic ecotope formation [6]. Similar processes exist in the metastasis of other digestive system tumors. miR-494, IL-1β and hGF promote hepatocellular carcinoma metastasis [76–78]; IL-8 and miR-21 promote gastric cancer metastasis [31, 79, 80];
Metabolic reprogramming
The organ tropism observed during tumor metastasis is closely associated with metabolic reprogramming in tumor cells, including alterations in glucose metabolism, lipid metabolism, and amino acid metabolism.
During liver metastasis in colorectal cancer, tumor cells exhibit enhanced dependence on amino acid metabolism. The glutamine metabolic pathway is significantly activated, with glutamine dehydrogenase (GDH) promoting metastasis and invasion of colorectal cancer cells through STAT3-mediated epithelial-mesenchymal transition (EMT) [81]. Additionally, lysine metabolism is involved in the colonization process, with its metabolic product acetyl-CoA activating the Wnt signaling pathway, while the generated glutamate helps maintain redox balance, thereby promoting liver colonization [82]. Furthermore, liver metastasis of colorectal cancer is associated with creatine levels. When creatine levels increase due to exogenous supplementation or endogenous synthesis, tumor metastatic capacity is enhanced [83].
In contrast, tumor cells in lung metastasis of colorectal cancer tend to activate lipid metabolic pathways. Related studies have shown that the expression of ATP-citrate lyase (ACLY) and stearoyl-CoA desaturase 1 (SCD1) is significantly upregulated in colorectal cancer cells. ACLY interacts with the CTNNB1 protein to stabilize it, and this complex may promote the transport of CTNNB1 from the cytoplasm to the nucleus, thereby enhancing the transcriptional activity of CTNNB1 and the migration and invasion capabilities of colon cancer cells, accelerating the process of colorectal cancer metastasis [84]. In addition, glycometabolic reprogramming plays a crucial role in the metastasis process in both liver and lung organs.
These findings not only reveal the key role of metabolic reprogramming in tumor metastasis organ selection but also provide a robust theoretical basis for future research into targeted therapeutic strategies tailored to different metastatic sites.
Immune system regulation
The immune system also plays an important role in organ tropism. Among these, neutrophils can systematically regulate the reprogramming of the pre-metastatic microenvironment, ultimately forming a pathological microenvironment that supports the colonization and proliferation of circulating tumor cells. Studies have shown that neutrophils can (i) degrade the extracellular matrix to promote tumor invasion, (ii) induce neovascularization to provide nutritional support, (iii) establish a pro-inflammatory environment and recruit immunomodulatory cells such as myeloid-derived suppressor cells (MDSCs) and tumor-associated macrophages, etc., through secretion of bioactive molecules, such as matrix metalloproteinases (MMPs), vascular endothelial growth factors (VEGFs), transforming growth factor β (TGF-βs), and interleukin 17 (IL-17s). In addition, MDSCs, tumor-associated macrophages, and other immunomodulatory cell populations play important biological roles, shaping multiple mechanisms for primary tumors and host factors to synergistically create suitable conditions for tumor metastasis (Table 2).
Anatomical and therapeutic factors
For colorectal cancer liver metastasis, the special anatomy of the portal vein also provides a shortcut for CTC convergent delivery from the intestine to the liver [6]which allows rapid encapsulation of CTC in the liver [120]. Interestingly, newer studies suggest that the effects produced by chemotherapeutic agents on the pre-metastatic microenvironment may promote the development of tumor metastasis. Significant reductions in the number of macrophages and T cells were observed in the livers of colorectal cancer patients treated with oxaliplatin chemotherapy. The number of macrophages and T cells was significantly reduced in the liver tissue of colorectal cancer patients who had received oxaliplatin, but the residual macrophages showed an immunosuppressive phenotype, an alteration that may play a role in synergistically promoting liver metastasis [121].
When CTCs leave the circulation, they may become disseminated tumor cells (DTCs) homing to new metastatic sites [7]. Interestingly, most CTCs enter a dormant state upon arrival at the target organ and, upon receiving metastatic site signals, reactivate uPAR and mitogenic signaling (ERBB2 or EGFR) to induce ERK activation and p38 inactivation, and ultimately switch to a value-added mode [85].
122]is constructed by enriching patients’ CTCs in peripheral blood and inoculating them into immune-deficient mice, which can expand primary tumor cells and generate clinically relevant xenograft tumors [49].
The first successful construction of the CDX model was achieved by researchers using the technique of immunodeficient mice undergoing intracardiac or tail vein injection of BC CTC-derived cell lines [123]. Currently, CDX has achieved relevant research results in breast cancer, prostate cancer, small-cell lung cancer, non-small-cell lung cancer, and melanoma [124]and has proven to be a reliable model for disease modeling and drug testing. However, the establishment and specific application of CDX models in digestive system tumors such as colorectal cancer and hepatocellular carcinoma still have a long way to go.
With technological advances in CTC isolation and enrichment strategies, we can expect higher CDX generation efficiency and wider clinical applications [125].
CTC-derived organoids(CDO)
CTC-derived organoids (CDO) can reflect the heterogeneity of primary tumors to a certain extent, and thus can be used as a prospective tool to dynamically monitor the evolutionary spectrum of tumor mutations [126]. It is safer to invade than patient-derived organoids (PDOs), exhibits a higher propensity to metastasize than primary-derived organoids [127]and may be able to better mimic tumor behavior and metastatic mechanisms [128]. CTC-derived colorectal cancer organoids have been successfully established using suspension culture methods, and for the first time, it has been demonstrated that CTCs isolated from CRC patients have a phenotype that expresses cancer stem cells, which can be used to determine drug sensitivity [129]providing a new idea for personalized medicine for patients with metastatic CRC.
130]. Its core definition is to capture tumor molecular information by detecting cancer-related cellular products such as circulating tumor cells (CTCs), circulating tumor DNA (ctDNA), cell-free circulating nucleic acids (cf. DNA/RNA), microRNAs (miRNA), long non-coding RNAs (lncRNAs), exosomes, and proteins in body fluids samples such as peripheral blood to capture tumor molecular information (Fig. 5).
Liquid biopsy. Liquid biopsy enables non-invasive tumor information capture by analyzing tumor-related molecular markers in body fluids such as peripheral blood, including circulating tumor cells (CTCs), circulating tumor DNA (ctDNA), cell-free circulating nucleic acids (cf. DNA/RNA), microRNAs long non-coding RNAs (lncRNAs), exosomes, and proteins, which provide the molecular basis for tumor dynamics monitoring and precision diagnosis and treatment.
As a non-invasive detection method, it can analyze CTCs or ctDNA in peripheral blood through dynamic repetitive sampling. With the breakthroughs in single-cell sequencing and microfluidic technology, liquid biopsy has been transformed from a theoretical concept into a key tool for clinical staging, early warning of recurrence, and evaluation of response to targeted therapies, which can realize the real-time tracking of the recurrence and metastasis process of tumors, as well as the dynamic evaluation of the This enables real-time tracking of tumor recurrence and metastasis and dynamic evaluation of treatment response, providing immediate molecular basis for individualized treatment.
As a “seed” in the process of tumor metastasis, CTC plays a pivotal role in the process of tumor metastasis, and naturally becomes a research hotspot in liquid biopsy. Moreover, CTC can provide tumor-specific information even after the primary tumor is removed [131].
Early detection and neoplasm staging
CTC is considered to be a key step in the initiation of tumor metastasis, so CTC may be one of the biomarkers for early detection of tumors. Early detection of tumors through CTC testing can help doctors and patients take more proactive treatment measures, and patients are expected to have a better prognosis.
For CRC, early diagnosis is essential to improve survival. Although colonoscopy is the “gold standard” for early diagnosis, patients are often resistant to it due to its invasive nature. Therefore, CTC, as a less invasive test, is more conducive to improving patient compliance.
Studies have shown that the sensitivity of preoperative CTC detection for CRC can reach 95.2% [132], which is very favorable for early detection of CRC. CTC counting by CellSearch in the clinic is currently approved by the FDA for the stratification of patients with tumors such as colorectal cancer [133]. In CRC, a lower threshold (≥ 3 CTC/7.5 mL) means a strong predictive value for PFS and OS [134]and corresponds to a higher risk of recurrence and significantly increased cancer-related death. However, the clinical utility of CTC testing in CRC remains to be further confirmed. Similarly in hepatocellular carcinoma, CTC, although highly correlated with TNM staging [135]can only be used as a complement to TNM staging in patients with HCC and cannot yet be used as an independent diagnostic basis. Similarly, relying on CTC count and phenotype alone is not sufficient to determine whether tumor cells will successfully colonize in distant organs, and a combination of other tests is still required.
Prognostic evaluation and recurrence monitoring
Nowadays, CTC counts and phenotypes have been shown to serve as biomarkers for metastatic risk stratification and prognostic assessment, and can guide clinicians in the optimization of individualized clinical decision-making through dynamic preoperative and/or postoperative monitoring [136, 137]. Studies have shown that patients with positive peripheral blood CTC have worse clinical outcomes [138, 139]. Studies have shown that HCC patients with high CTC counts are significantly associated with poor prognosis after radical resection [140]; the persistent presence of CTC postoperatively is a predictor of early recurrence in patients with colon cancer [141]. In addition to CTC count alone, different CTC subtypes are also valuable for research. CTC can be categorized into epithelial CTC, epithelial/mesenchymal CTC, and mesenchymal CTC subtypes based on EpCAM, CK8/18/19, vimentin, and twist. It has been shown that in colorectal cancer, total CTC and mesenchymal CTC are significantly associated with advanced disease stage [142] and have a higher sensitivity for predicting lymph node involvement and/or distant metastasis [143]; similarly, in hepatocellular carcinoma it has been shown that mesenchymal CTC is associated with early recurrence [144]; EpCAM + CTC can be used to monitor treatment efficacy while also being a target for the treatment of HCC recurrence [145]. The above findings suggest that CTC counts may be useful as an indicator for determining the prognosis of patients with tumors and that specific CTC phenotypes may be relatively more valuable. However, some studies have also indicated that CTC trajectories are more appropriate prognostic indicators compared to CTC counts at baseline. Using serial CTC assessments to define CTC trajectories might optimize risk stratification and help monitor mCRC treatment response [146].
Treatment guidelines
With the in-depth exploration of CTC-mediated metastasis mechanisms, how CTCs guide clinical treatment has attracted more attention.
Personalized treatment based on CTC count guidance
CTC may be an indicator of patient selection for individualized treatment modalities. Studies have shown that CTC counts can screen patients suitable for receiving intensive first-line FOLFOXIRI plus bevacizumab [105]. Another study indicated that preoperative peripheral blood CTCs counts in patients with pancreatobiliary cancer may be a biomarker to predict positive surgical margins [147].
Targeted CTC intervention
Since CTCs are a key factor in mediating tumor metastasis, some scholars have asked whether the development of tumor metastasis can be terminated if CTC is eliminated, thus prolonging the survival of tumor patients. It has been demonstrated in a CTC-loaded mouse model that selective elimination of CTC can effectively inhibit tumor metastasis and significantly prolong the survival of loaded mice [148].
Targeted at immune checkpoints
In recent years, with the continuous exploration of tumor immune escape mechanisms, immune checkpoint blockade (ICB) has made unprecedented progress in cancer treatment. A study on pancreatic ductal adenocarcinoma (PDAC) [149] analyzed the transcriptomes of PDAC patients’ circulating tumor cells (CTCs), primary tumors, and metastatic tumors at the single-cell level, revealing that CTCs can activate HLA-E expression by taking up RGS18 from platelets, thereby binding to CD94-NKG2A on the surface of natural killer (NK) cells and inhibiting their cytotoxic function. In vitro and in vivo experiments have confirmed that blocking the HLA-E: CD94-NKG2A interaction can effectively restore the immune surveillance function of NK cells and their cytotoxic activity against CTCs, and inhibit tumor metastasis mediated by CTCs. Another study [42] about HCC indicated that circulating tumor cells (CTCs) primarily evade NK cell-mediated innate immune surveillance through the immune checkpoint CD155-TIGIT. After blocking the CD155-TIGIT interaction using TIGIT antibodies, NK cell immune surveillance and attack against CTCs were restored, and tumor metastasis was significantly reduced.
In addition, gut microbiota metabolites also play a role in enhancing the anti-tumour capacity of immune cells. Studies have shown that butyrate can enhance the antitumor immunity of cytotoxic CD8 + T cells [150] and improve the efficacy of anti-PD-1 immunotherapy [151]making it a promising biomarker for antitumor immunotherapy. The rapid progress of such research provides new immunotherapy strategies for preventing tumor metastasis.
Targeted at CTC-neutrophil clusters
Previous studies have also shown that neutrophils can accelerate metastatic seeding through direct interaction with CTCs [44]suggesting that CTC-neutrophil clusters could be a new therapeutic target. Current therapeutic approaches targeting CTC-neutrophil clusters primarily focus on CTCs, neutrophils, and NETs [152]including but not limited to nanodrugs that precisely target CTCs to induce apoptosis [153]nanoparticles that inhibit NET formation [154]inhibiting neutrophil migration [155]and blocking the formation of CTC-neutrophil clusters [156]among others.
In addition, many research results have shown that the number and metastatic potential of CTC fluctuate with circadian rhythms [157, 158]and such a cyclic pattern of change may contribute to the further development of detection methods and selection of therapeutic strategies. At the same time, in addition to the many factors that affect the “seed” itself, the homeostatic factors that create the “soil” environment should also be emphasized [67]. CTC is a promising therapeutic target in the field of tumor metastasis research, which is expected to help tumor patients reduce the risk of metastasis.
CTC enrichment strategies. Enrichment based on biochemical characterization includes methods such as positive enrichment methods, negative enrichment, and nanotechnology. Enrichment based on physical properties includes methods such as filtration, density gradient centrifugation and microfluidic chips
Enrichment strategies based on biochemical characterization
Enrichment based on biochemical characterization mainly screens out CTCs based on proteins specifically expressed on the cell surface, and includes methods such as positive enrichment (capture of CTCs), negative enrichment (depletion of background blood cells [159]), and flow cytometry.
CellSearch represents an application of the positive enrichment method, in which anti-epithelial cell adhesion molecule (EpCAM) specific antibodies are coated on magnetic beads and used to capture CTCs.CellSearch is licensed by the Food and Drug Administration (FDA) for the detection of CTCs in CRCs. the gold standard [160]. In addition to CellSerach, CTC microarrays (CTCs-chip) and the MACS® Cell Separation magnetic cell sorting system are also positive enrichment methods. The advantage of these methods is the high purity of the captured CTCs. The disadvantage is that only epithelial (E-type) CTCs and hybrid (H-type) CTCs can be captured in blood, but not EpCAM-negative mesenchymal (M-type) CTCs; in addition, epithelial markers may be down-regulated or lost in the EMT process, which may lead to false-negative results [161].
Negative enrichment, also known as leukocyte removal. It uses specific antibodies CD45, CD14, etc. to bind to leukocytes, thus indirectly enriching CTC precipitation. A typical example is the Cyttel/Cytelligen detection system. Compared with the positive enrichment method, the negative enrichment method can detect EpCAM-negative and CK-negative mesenchymal (M-type) CTCs; however, it requires more steps in sample processing, and the already scarce CTCs are easily lost.
Furthermore, Racila et al. performed the first clinical trial combining DAPI-positive, CK-positive, and CD45-negative cell isolation techniques with flow cytometry in 1998 [162].Terstappen integrated EpCAM assay for CTC with flow cytometry to combine the two [163].
Enrichment strategies based on physical characterization
Enrichment based on physical properties is mainly based on the diameter, density and other characteristics of CTCs to screen out these cells, including filtration (CTC volume is larger than blood cells), density gradient centrifugation (CTCs have different densities from leukocytes) and microfluidic chips.
With technological innovation and development, traditional methods have been complemented by more advanced technologies such as microfluidics, next generation sequencing (NGS) and single cell RNA sequencing (scRNA-seq). Microfluidic devices have gained attention for their ability to capture CTC based on physical properties such as size and deformability. Bao-Caamano et al. [164] analyzed the epigenomic changes of CTC in CRC patients using a microfluidic system. Their study revealed a unique DNA methylation program in metastatic CTC, highlighting the potential of microfluidics in understanding CTC biology.
Microfluidic devices have gained attention for their ability to capture CTCs based on physical properties such as size and deformability utilized a microfluidic system to analyze the epigenomic changes in CTCs from CRC patients. Their study revealed a unique DNA methylation program in metastasis-competent CTCs, highlighting the potential of microfluidic technologies for understanding CTC biology.
Besides, microfluidic rare-cell capture technology (microfluidic rare-cell capture technology) can provide technical support for early cancer detection and monitoring [165, 166]. In addition, nanotechnology, which is in full swing, is dedicated to improving the sensitivity and specificity of CTC detection [160]. Xenotransplantation into mice can enrich the number of CTCs while promoting new animal studies [167].
These techniques have improved the sensitivity and specificity of CTC detection, allowing for a more comprehensive understanding of its biological properties and giving new impetus to CTC-related research. However, due to the extremely low concentration of CTC, its short half-life in the circulation (1–2.5 h) [168]and its high heterogeneity (lack of tumor-specific markers), it is still difficult to accurately capture CTC, which is a great challenge in clinical practice [169]and greatly reduces the value of its practical application [170].
Single-cell technology applications
With the rapid development of single-cell technology, its exploration and research in the field of circulating tumor cell (CTC) studies have also become increasingly in-depth.
Uniform Chromosome Conformation Capture (Uni-C) developed by researchers can simultaneously analyses genomic structural variations (SV, CNV, ecDNA), small-scale mutations (SNP/INDEL), and chromatin three-dimensional conformation at the single-cell level. In Pancreatic cancer CTC research, it was found that only 7 cells are required to reconstruct 89% of SNPs/INDELs and 75% of structural variations, while also resolving the molecular structure of ecDNA and cell cycle-dependent chromatin conformation differences [171]which can be used for neoantigen prediction and personalized immunotherapy design. Single-cell metabolomics technology platforms can efficiently combine CTC capture with single-cell metabolic analysis. By predicting tumor metastasis risk through metabolic fingerprints (e.g., phenylalanine and arginine pathways), the accuracy rate reaches 74–92% [172]. Additionally, single-cell RNA sequencing (scRNA-seq) is currently used in the CTC field to analyze CTC transcriptional heterogeneity and reveal the expression dynamics of metastasis-related genes (such as EMT markers and drug resistance genes). Studies have shown that CTCs sorted by LIFT-AMFS have been validated by scRNA-seq for genomic integrity and transcriptional reliability [173].
The core challenges of current CTC single-cell technologies lie in the efficient capture of rare cells and the integration of multidimensional data. We speculate that future trends may be related to automated platforms that directly couple microfluidic enrichment with single-cell sequencing, combining organoid culture or animal model validation of CTC metastatic potential, and individualized neoantigen therapy for clinical translation.
In terms of biological mechanisms, more emphasis should be placed on combining with single-cell genomics [176] (genome, transcriptome, proteome) and organoid co-culture models to elucidate the clonal evolution process (e.g., screening of the most invasive subpopulations), dormancy regulation, molecular mechanisms of intravasation/extravasation, and their interactions with the microenvironment (shear force, exosomes, immune cells), so as to provide targets for anti-metastatic therapy.
It is worth noting that single-cell technology plays a pivotal role at both the technical and mechanistic levels. The rapid development of single-cell technology has provided unprecedented technical support for in-depth research into the molecular characteristics of circulating tumor cells (CTCs). In recent years, with the mature application of high-precision analytical technologies such as single-cell sequencing and single-cell proteomics, researchers have been able to analyse the heterogeneous characteristics of CTCs from multiple dimensions, including the genome, transcriptome, epigenome, and proteome. These technological advancements not only hold promise for revealing the molecular classification mechanisms of CTC subpopulations but also provide a solid scientific basis for establishing a precise classification system for CTCs based on molecular characteristics. It is worth noting that current research on tumor organ tropism indicates that this process is closely related to key factors such as chemokine networks, vascular endothelial characteristics, and immune cell infiltration in the tumor microenvironment. As the ‘seeds’ of distant tumor metastasis, the chemokine receptors, integrin family molecules, and metabolic reprogramming characteristics specifically expressed on the surface of CTCs may be mechanistically associated with their selective settlement in target organs. Therefore, future research may focus on integrating single-cell multi-omics technologies and in vitro/in vivo metastasis models to systematically elucidate the regulatory relationships between CTC molecular profiles and specific metastatic target organs. This could have significant implications for the development of early intervention and precision treatment strategies for tumor metastasis.
In terms of clinical application translation, there are also future breakthroughs that are worthwhile, including (i) Early diagnosis and dynamic monitoring: combining other liquid biopsy components such as cfDNA and exosomes to improve the early cancer detection rate and early warning of micrometastasis [177]; (ii) Prognosis and treatment guidance: constructing predictive models and real-time drug sensitivity detection by optimizing CTC in vitro culture; (iii) Intervention strategy development: targeting key factors in the key steps of CTC dissemination, and exploring corresponding interventions by considering the mutational background.
However, the standardization and validation of CTC research is still the core bottleneck, perhaps through the unification of cross-platform technological processes as well as the conduct of large-scale interventional clinical trials to validate the degree of its universality in different cancer types [178]and the optimization of the detection time window based on the CTC biorhythms, which will ultimately achieve the leap from basic research to precision oncology medicine with CTC.
XAH24007) the National Natural Science Foundation of China (82303923), Guangdong Basic and Applied Basic Research Foundation (2024A1515013156).